Recombinant BjADAT2 and BjADAT3 were individually expressed in E. coli and purified, and their enzyme activities were tested by in vitro tRNA deamination assay (Supplementary Fig. 4). Neither BjADAT2 nor BjADAT3 showed any A-to-I editing activity, but BjADAT2, in complex with BjADAT3, could perform A-to-I editing of amphioxus tRNAVal(AAC), and the apparent first-order deamination rate constant (kapp) of tRNA deamination was 0.0287 ± 0.00219 min−1. Because the optimal RNA substrate for ADAT2 was the adenosines on the anticodon loop structure of tRNA, we wondered if BjADAT2 could mediate DNA deamination in a structure-specific fashion. Thus, we compared the deamination efficiency of BjADAT2 on DNA hairpin structure substrates (i.e., hpDNA-A and hpDNA-C, containing a single adenosine or cytidine in the loop region, respectively) and single-stranded linear structure substrates (i.e., ssDNA-A and ssDNA-C, containing a single adenosine or cytidine in the substrates, respectively). As shown in Fig. 3, both adenosine deamination of hpDNA-A and cytidine deamination of hpDNA-C were clearly observed under BjADAT2 treatment, and the change of the hairpin substrates to linear substrates resulted in about a threefold decrease in adenosine-and cytidine-deamination ratios. By contrast, no product band was seen in the lane of hpDNA-G or hpDNA-T treated with BjADAT2. Moreover, the BjADAT2-E58A protein purified in the same way as BjADAT2 showed no deamination activity (Supplementary Fig. 5f, g), ruling out the possibility that the observed deamination activity of BjADAT2 arose from a contaminant in the recombinant protein samples. In addition, BjADAT2-mediated adenosine-and cytidine deamination was inhibited by deoxycoformycin (DCF), an adenosine deaminase specific inhibitor, or tetrahydrouridine (THU), a cytidine deaminase specific inhibitor, or 1,10-o-phenanthroline, a zinc chelator. These indicated that BjADAT2 deaminated adenosine and cytidine in a zinc-dependent manner.
The kapp of BjADAT2-induced hpDNA-A deamination (0.00206 ± 0.00019 min−1) was comparable with that of ABE7.10-induced DNA adenosine deamination (0.0010 ± 0.00030 min−1)22. We compared the kinetics of BjADAT2-induced DNA deamination of the hairpin substrates and the linear substrates. The kapp was sixfold higher for the hpDNA-A substrate than for the ssDNA-A substrate (0.00032 ± 0.00004 min−1). Similarly, the kapp was 24-fold higher for hpDNA-C (0.00801 ± 0.00037 min−1) than for ssDNA-C (0.00033 ± 0.000026 min−1) (Fig. 3e). These data together indicated that BjADAT2 preferentially deaminates adenosines and cytidines in the hairpin loop of substrates.
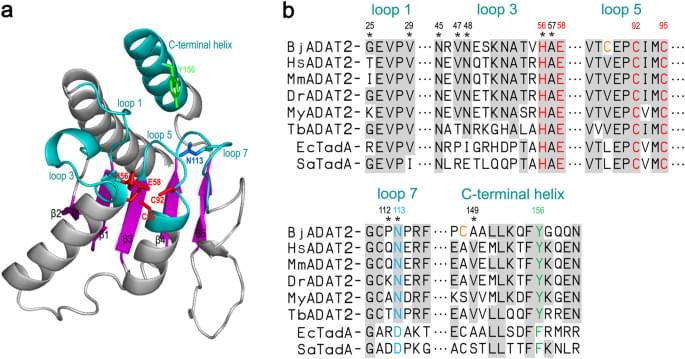
It was known that ADAT3, as a non-catalytic subunit, served a structural role in the adenosine deamination of tRNA10,23. To date, only trypanosome ADAT2 was shown to be able to deaminate cytosines of DNA in the presence of ADAT3 in vitro6, but the specific role of the ADAT3 subunit in this DNA editing reaction was unclear. We demonstrated here that BjADAT3, though lacking catalytic function, could enhance the deamination activity of BjADAT2 toward hairpin structure substrates, including hpDNA-A and hpDNA-C (Fig. 3a, b). This was also supported by the results of kinetic analysis (Fig. 3e), which showed that the kapp of hpDNA-A deamination of BjADAT2/BjADAT3 complex (0.01092 ± 0.00062 min−1) was 5.3-fold higher than that of BjADAT2 alone, and the kapp of hpDNA-C deamination by the complex (0.01066 ± 0.00083 min−1) 1.3-fold higher than that of BjADAT2 alone. On the contrary, the kapp of ssDNA-A deamination of BjADAT2/BjADAT3 complex (0.00016 ± 0.